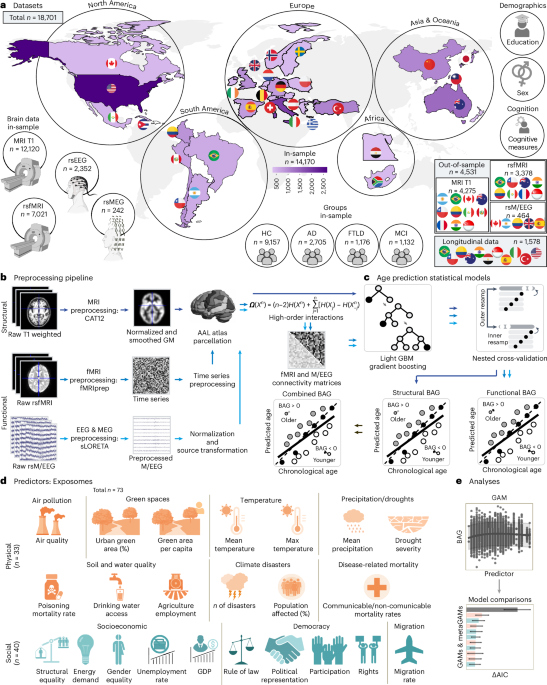
Abstract
The physical and social exposome affects human aging, and brain clocks may track its effects. However, most studies neglect multidomain exposures (physical, social and political) across diverse settings globally and their associations with brain aging. In this study, we characterized the associations between 73 country-level physical and social exposomal factors and multimodal brain age in 18,701 participants from 34 countries (healthy individuals and those with Alzheimer’s disease, frontotemporal lobar degeneration or mild cognitive impairment). Exposome effects were assessed using generalized additive models and meta-analytic frameworks. Aggregated exposome models explained up to 15.5-fold more variance than individual exposures (delta Akaike information criterion (ΔAIC): 2,034–3,127). Physical exposome was primarily associated with accelerated structural brain aging (limbic, subcortical and cerebellar regions), whereas social exposome was more strongly associated with functional brain aging (frontotemporal and limbic networks). Exposome burden accounted for 3.3−9.1-fold higher risk of accelerated aging, exceeding effects of clinical diagnoses. Findings were out-of-sample validated in cross-sectional and longitudinal designs, remained consistent across clinical subgroups and persisted after adjustment for demographics, age correction bias, cognition, scanner type and data quality. The exposome accelerates brain aging in health and disease, underscoring the need to address physical, social and political inequities.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others
Data availability
Preprocessed BAGs and exposome values are provided online (https://osf.io/yagdh The brain datasets used in this study are available from different sources (Supplementary Table 35). The MRI T1, rsfMRI and rsM/EEG datasets comprise sources: (1) currently publicly available for direct download after registration, (2) available after contacting the researcher or (3) accessible after institutional review board approval of formal data-sharing agreement in a process that can last up to 12 weeks, depending on the nature of the request and compliance with institutional guidelines. The exposome datasets come from sources that are either publicly available for direct download or accessible upon reasonable request (Supplementary Tables 1 and 2). Individual-level neuroimaging and clinical data are not publicly shared due to ethical and institutional restrictions imposed by the original data providers. External requests for individual-level data must be directed to the corresponding data-owning institutions listed in Supplementary Table 35 and will be evaluated according to their local governance, institutional review board requirements and data-sharing policies; response times typically range from 4 weeks to 12 weeks.
Code availability
Preprocessing and analysis codes are freely available on the Open Science Foundation repository: https://osf.io/yagdh A README file has been added to the repository, which includes (1) a project overview, (2) data description and input/output formats, (3) setup and dependencies, (4) step-by-step instructions for running preprocessing and analysis scripts, (5) workflow for reproducing figures and tables and (6) directory structure with script functions.
References
Ibanez, A. et al. Computational whole-body-exposome models for global precision brain health. Nat. Commun. 16, 11078 (2025).
Miller, G. W. Banbury Exposomics Consortium. Integrating exposomics into biomedicine. Science 388, 356–358 (2025).
Ibanez, A. et al. Neuroecological links of the exposome and One Health. Neuron 112, 1905–1910 (2024).
Migeot, J. et al. Social exposome and brain health outcomes of dementia across Latin America. Nat. Commun. 16, 8196 (2025).
Belsky, D. W. & Baccarelli, A. A. To promote healthy aging, focus on the environment. Nat. Aging 3, 1334–1344 (2023).
Argentieri, M. A. et al. Integrating the environmental and genetic architectures of aging and mortality. Nat. Med. 31, 1016−1025 (2025).
Hernandez, H. et al. The exposome of healthy and accelerated aging across 40 countries. Nat. Med. 31, 3089–3100 (2025).
Moguilner, S. et al. Brain clocks capture diversity and disparities in aging and dementia across geographically diverse populations. Nat. Med. 30, 3646–3657 (2024).
Bittner, N. et al. When your brain looks older than expected: combined lifestyle risk and BrainAGE. Brain Struct. Funct. 226, 621–645 (2021).
Wu, Y. et al. Flooding exposure accelerated biological aging: a population-based study in the UK. Environ. Res. Lett. 19, 074051 (2024).
Gong, J. & Zaninotto, P. Integrating exposome into lifecourse understanding of cognitive ageing and dementia: current evidence, methodological challenges, and future directions. Int. J. Environ. Res. Public Health 22, 815 (2025).
Pu, F. et al. Heterogeneous associations of multiplexed environmental factors and multidimensional aging metrics. Nat. Commun. 15, 4921 (2024).
Ruas, M. V. et al. Large-scale georeferenced neuroimaging and psychometry data link the urban environmental exposome with brain health. J. Environ. Res. 267, 120632 (2025).
Sakowski, S. A., Koubek, E. J., Chen, K. S., Goutman, S. A. & Feldman, E. L. Role of the exposome in neurodegenerative disease: recent insights and future directions. Ann. Neurol. 95, 635–652 (2024).
Ibanez, A., Kringelbach, M. L. & Deco, G. A synergetic turn in cognitive neuroscience of brain diseases. Trends Cogn. Sci. 28, 319−338 (2024).
Pandics, T. et al. Exposome and unhealthy aging: environmental drivers from air pollution to occupational exposures. Geroscience 45, 3381–3408 (2023).
Xiang, W. et al. Chronic high temperature exposure, brain structure, and mental health: cross-sectional and prospective studies. Environ. Res. 264, 120348 (2025).
Bao, Y. et al. Water quality and neurodegenerative disease risk in the middle-aged and elderly population. Ecotoxicol. Environ. Saf. 289, 117647 (2025).
Hunter, R. F. et al. Advancing urban green and blue space contributions to public health. Lancet Public Health 8, e735–e742 (2023).
Baez, S. et al. Structural inequality and temporal brain dynamics across diverse samples. Clin. Transl. Med. 14, e70032 (2024).
Legaz, A. et al. Structural inequality linked to brain volume and network dynamics in aging and dementia across the Americas. Nat. Aging 5, 259–274 (2025).
Zugman, A. et al. Country-level gender inequality is associated with structural differences in the brains of women and men. Proc. Natl Acad. Sci. USA 120, e2218782120 (2023).
Hernandez, H. & Ibanez, A. Expotype−phenotype resilience and multimodal aging clocks. Clin. Transl. Med. 16, e70558 (2026).
Barnes, L. L. Alzheimer disease in African American individuals: increased incidence or not enough data? Nat. Rev. Neurol. 18, 56–62 (2022).
Herzog, R. et al. Genuine high-order interactions in brain networks and neurodegeneration. Neurobiol. Dis. 175, 105918 (2022).
Coronel-Oliveros, C. et al. Diversity-sensitive brain clocks linked to biophysical mechanisms in aging and dementia. Nat. Ment. Health 3, 1214–1229 (2025).
Ke, G. et al. LightGBM: a highly efficient gradient boosting decision tree. In NIPSʼ17: Proceedings of the 31st International Conference on Neural Information Processing Systems (eds von Luxberg, U. & Guyon, Y.) 3149–3157 (Curran Associates, 2017).
Schulz, M.-A., Siegel, N. T. & Ritter, K. Brain-age models with lower age prediction accuracy have higher sensitivity for disease detection. PLoS Biol. 23, e3003451 (2025).
Butler, E. R. et al. Pitfalls in brain age analyses. Hum. Brain Mapp. 42, 4092–4101 (2021).
Burnham, K. P. & Anderson, D. R. Multimodel inference: understanding AIC and BIC in model selection. Sociol. Methods Res. 33, 261–304 (2004).
Mendenhall, E., Kohrt, B. A., Logie, C. H. & Tsai, A. C. Syndemics and clinical science. Nat. Med. 28, 1359–1362 (2022).
Liu, F. et al. Environmental neuroscience linking exposome to brain structure and function underlying cognition and behavior. Mol. Psychiatry 28, 17–27 (2023).
Cristaldi, A. et al. Possible association between PM2.5 and neurodegenerative diseases: a systematic review. Environ. Res. 208, 112581 (2022).
Qin, S. J. et al. Neurotoxicity of fine and ultrafine particulate matter: a comprehensive review using a toxicity pathway-oriented adverse outcome pathway framework. Sci. Total Environ. 947, 174450 (2024).
Vanbrabant, K. et al. Accumulation of ambient black carbon particles within key memory-related brain regions. JAMA Netw. Open 7, e245678 (2024).
Li, K. et al. Association of environmental exposome and cognitive function among older adults with and without preclinical Alzheimer’s disease. Alzheimers Dement. 21, e70373 (2025).
McEwen, C. A. Connecting the biology of stress, allostatic load and epigenetics to social structures and processes. Neurobiol. Stress 17, 100426 (2022).
Martin, C. L., Ghastine, L., Lodge, E. K., Dhingra, R. & Ward-Caviness, C. K. Understanding health inequalities through the lens of social epigenetics. Ann. Rev. Public Health 43, 235–254 (2022).
Santamaría-García, H. et al. Allostatic interoceptive overload across psychiatric and neurological conditions. Biol. Psychiatry 97, 28–40 (2025).
Chen, B. et al. Contrasting inequality in human exposure to greenspace between cities of Global North and Global South. Nat. Commun. 13, 4636 (2022).
Iaccarino, L. et al. Association between ambient air pollution and amyloid positron emission tomography positivity in older adults with cognitive impairment. JAMA Neurol. 78, 197–207 (2021).
Da Ros, L. U. et al. Social and health disparities associated with healthy brain ageing in Brazil and in other Latin American countries. Lancet Glob. Health 13, e277–e284 (2025).
Legaz, A., Baez, S. & Ibañez, A. Unequal burdens: how structural socioeconomic inequality shapes brain health in aging and dementia. Neuroscience 569, 245–247 (2025).
Daly, T., Slachevsky, A., Trépel, D., Walsh, S. & Ibáñez, A. Accounting for inequality in dementia prevention programmes. Bull. World Health Organ. 104, 127−130 (2025).
Migeot, J. et al. Dementia prevention requires moving beyond individual choice: the costs of effort and time intersect with social determinants of health. Neuroscience 589, 256−267 (2025).
Livingston, G. et al. Dementia prevention, intervention, and care: 2024 report of the Lancet standing Commission. Lancet 404, 572–628 (2024).
Hartung, T. How AI can deliver the Human Exposome Project. Nat. Med. 31, 1738 (2025).
Ibanez, A. & Farombi, T. Biological imprint of education and rights-based brain capital. Alzheimers Dement. 21, e70222 (2025).
Mostert, C. M. et al. Broadening dementia risk models: building on the 2024 Lancet Commission report for a more inclusive global framework. EBioMedicine 120, 105950 (2025).
Peterson, R. L., Carvajal, S. C., McGuire, L. C., Fain, M. J. & Bell, M. L. State inequality, socioeconomic position and subjective cognitive decline in the United States. SSM Popul. Health 7, 100357 (2019).
González-Rodríguez, P., Füllgrabe, J. & Joseph, B. The hunger strikes back: an epigenetic memory for autophagy. Cell Death Differ. 30, 1404–1415 (2023).
Baez, S., Nabil, Y. & Ibanez, A. Childhood adversities and the exposome in dementia risk and brain health. Curr. Opin. Psychiatry 39, 136–150 (2026).
Hossin, M. Z. International migration and health: it is time to go beyond conventional theoretical frameworks. BMJ Glob. Health 5, e001938 (2020).
Coronel-Oliveros, C. et al. Creative experiences and brain clocks. Nat. Commun. 16, 8336 (2025).
Beck, D. et al. White matter microstructure across the adult lifespan: a mixed longitudinal and cross-sectional study using advanced diffusion models and brain-age prediction. Neuroimage 224, 117441 (2021).
Santamaria-Garcia, H. et al. Factors associated with healthy aging in Latin American populations. Nat. Med. 29, 2248–2258 (2023).
Amoruso, L. et al. Multilingualism protects against accelerated aging in cross-sectional and longitudinal analyses of 27 European countries. Nat. Aging 5, 2340–2354 (2025).
Ibanez, A. & Slachevsky, A. Environmental–genetic interactions in ageing and dementia across Latin America. Nat. Rev. Neurol. 20, 571–572 (2024).
Udeh-Momoh, C. T. Resilience and brain health in global populations. Nat. Med. 31, 2518–2531 (2025).
Gonzalez-Gomez, R. et al. Qualitative and quantitative educational disparities and brain signatures in healthy aging and dementia across global settings. EClinicalMedicine 82, 103187 (2025).
McKhann, G. M. et al. The diagnosis of dementia due to Alzheimer’s disease: recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 7, 263–269 (2011).
Gorno-Tempini, M. L. et al. Classification of primary progressive aphasia and its variants. Neurology 76, 1006–1014 (2011).
Rascovsky, K. et al. Sensitivity of revised diagnostic criteria for the behavioural variant of frontotemporal dementia. Brain 134, 2456–2477 (2011).
Petersen, R. C. et al. Mild cognitive impairment: clinical characterization and outcome. Arch. Neurol. 56, 303–308 (1999).
Mohammadi-Nejad, A.-R. et al. Accelerated brain ageing during the COVID-19 pandemic. Nat. Commun. 16, 6411 (2025).
Prado, P. et al. Harmonized multi-metric and multi-centric assessment of EEG source space connectivity for dementia characterization. Alzheimers Dement. (Amst.) 15, e12455 (2023).
Gonzalez-Gomez, R. et al. Educational disparities in brain health and dementia across Latin America and the United States. Alzheimers Dement. 20, 5912–5925 (2024).
Varma, S. & Simon, R. Bias in error estimation when using cross-validation for model selection. BMC Bioinformatics 7, 91 (2006).
Chen, H., Lundberg, S. M. & Lee, S.-I. Explaining a series of models by propagating Shapley values. Nat. Commun. 13, 4512 (2022).
Zhao, L. et al. Quantitative signal quality assessment for large-scale continuous scalp electroencephalography from a big data perspective. Physiol. Meas. 44, 035009 (2023).
Murtaugh, P. A. In defense of P values. Ecology 95, 611–617 (2014).
Ho, D. E., Imai, K., King, G. & Stuart, E. A. Matching as nonparametric preprocessing for reducing model dependence in parametric causal inference. Political Analysis 15, 199–236 (2007).
Li, Y., Xu, L. & Yamanishi, K. GMMDA: Gaussian mixture modeling of graph in latent space for graph data augmentation. Knowl. Inf. Syst. 66, 7667–7695 (2024).
Tian, Y. E. et al. Heterogeneous aging across multiple organ systems and prediction of chronic disease and mortality. Nat. Med. 29, 1221–1231 (2023).
Acknowledgements
A.L. is supported by a ReDLat-NIH postdoctoral position and an Atlantic Fellowship from the Global Brain Health Institute (GBHI). H.H. is supported by the Davos Alzheimer’s Collaborative. F.F. is supported by an Alzheimer’s Association Research Fellowship Grant (AARF-21-848281). S.A. is supported by IN212624, PAPIIT-DGAPA-UNAM IN208622 y SECIHTI CF-2019/6390. L.A. is supported by the European Union’s Horizon 2020 research and innovation program under Marie Sklodowska-Curie grant agreement number 101025814. M.I.B. is supported by National Institute on Drug Abuse 75N95022C00031: Genetics of Alzheimer’s Disease and Related Dementias in Latin America. M.I.B. is also co-investigator for SG-20-725707-ReDLat, Alzheimer’s Association, Multi-partner Consortium for Dementia Research in Latin America. O.C. is supported by the French government under management of Agence Nationale de la Recherche as part of the ‘France 2030’ program (reference ANR-23-IACL-0008, project PRAIRIE-PSAI), as part of the ‘Investissements d’avenir’ program (reference ANR-19-P3IA-0001, project PRAIRIE 3IA Institute and reference ANR-10-IAIHU-06, project Agence Nationale de la Recherche-10-IA Institut Hospitalo-Universitaire-6), and by the European Union’s Horizon Europe Framework Program (grant number 101136607, project CLARA). N.C. is supported by ReDLat-NIH (AG057234). S.D. is supported by a grant overseen by the French National Research Agency (ANR) as part of the France 2030 ‘Investment for the Future Programme’ through the SHIVA project (ANR-18-RHUS-0002) and through the Precision and Global Vascular Brain Health Institute, funded by the France 2030 IHU3 initiative (ANR-23-IAHU-0001). Computations were performed at the Bordeaux Bioinformatics Center (CBiB) and the CREDIM computer resources, University of Bordeaux. Funding support for additional computer resources has been provided to S.D. by the Fondation Claude Pompidou. C.C. is supported by the National Medical Research Council of Singapore. The i-Share study has received funding from the French National Agency (Agence Nationale de la Recherche (ANR)), via the Investment for the Future program (grant no. ANR-10-COHO-05 and no. ANR-18-RHUS-0002) and from the University of Bordeaux Initiative of Excellence (IdEX). P.D. is supported by Polish National Science Center (NCN) grant number 2018/31/N/HS6/03551. C.J. is supported by UNQ, Proyecto de I+D Res.745/25. E.K. is supported by the MES-CoBraD Project EU Horizon 2020 grant (ID: 965422) to the Neurological Institute of Athens. J.L. is supported by the US National Institute on Aging (NIA), National Institutes of Health (NIH) (R01AG051125 and U01AG064948). L.A. is supported by the Office of the Director, NIH, under award numbers S10OD032285, R01AG080473, RF1AG087965 and RF1AG088003. (The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.) J.H.Z. is supported by the Singapore National Medical Research Council, grant numbers NMRC/OFLCG19May-0035, HLCA23Feb-0004 and OFIRG24Jul-0049. S.L. is supported by the ‘BE FOR ERC 2023’ fellowship from Sapienza University of Rome. P.M. is supported by the Belmon Forum. G.N. would like to extend her gratitude to V. Aliji from the University Institute of Radiology-Skopje, North Macedonia, and the Neurophysiology team from the University Clinic of Neurology, Skopje, North Macedonia, for their cooperation. P.O. is supported by Proyecto Fondecyt 1241469, ANID, Chile, and Proyecto AFB240002, ANID, Chile. M.A.P. is supported by a BrainLat Seed Grant (BL-SRGP2020-020) awarded to M.A.P. and A.I. E.d.P.F.R. is supported by the NIA of the NIH (R21AG069252), the Alzheimer’s Association (GBHI_ALZ-18-534892) and the World Federation of Neurology. G.A.S.J. is supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq grant numbers 573974/2008-0 and 465550/2014-2); Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP grant numbers 2008/57896-8, 2013/08531-5, 2014/50917-0, 2020/06172-1, 2021/05332-8 and 2021/12901-9); the European Research Council (ERC grant numbers 337673 and 101057390); the UK Medical Research Council (MRC grant number MR/R022763/1); Ministério da Saúde (Decit/SECTICS/MS grant number 888379/2019, Portaria no. 1.949, 04/08/2020); and Banco Industrial do Brasil S/A (CISM grant). A.S. is supported by ANID/Fondap/15150012, ANID/Fondecyt/1231839, ANID/FONDEF/ID22I10251 and ANID/Proyectos de Exploración 13220082 and ReDLat (supported by NIA, NIH (R01 AG057234), the Alzheimer’s Association (SG-20-725707), the Tau Consortium and the GBHI) and by Alzheimer’s Association GBHI ALZ UK-20-639295. L.T.T. is supported by the NIH (ReDLat). J.C. is supported by ANID (FONDECYT Postdoctorado no. 3240042 and Proyectos de Exploración no. 13240170). R.H. is supported by the Spanish Ministerio de Ciencia, Innovación y Universidades (MICIU/AEI/10.13039/501100011033) through María de Maeztu project CEX2021-001164-M. G.F.M. is supported by ANID (Fondecyt Postdoctoral no. 3250669). A.M.G. is partially supported by the NIA of the NIH (R01AG075775, R01AG083799 and 2P01AG019724); ANID (FONDECYT Regular 1250317 and 1250091); DICYT-USACH (032351GDAS); Agencia Nacional de Promoción Científica y Tecnológica (01-PICTE-2022-05-00103); and the ReDLat, which is supported by the Fogarty International Center and the NIH, NIA (R01AG057234, R01AG075775, R01AG21051 and CARDA-NIH), the Alzheimer’s Association (SG-20-725707), the Rainwater Charitable Foundation’s Tau Consortium, the Bluefield Project to Cure Frontotemporal Dementia and the GBHI. I.G.-C. is supported by an AFTD Pathways for Hope Pilot Grant (2025-002). R.L.-R. is supported by a National Health and Medical Research Council (NHMRC) Investigator Grant (GNT2010064). M.E.L. is supported by I+D+i project PID2022-141524NA-I00, funded by MCIN/AEI/10.13039/501100011033. V.M. is supported by ANID/FONDECYT de Iniciación 11251578 and ANID/FONDECYT Exploración 13240170. D.R.G. is supported by Fellowship 1003248 from Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI). H.S.G. is supported by an NIH R01 grant (‘Social epigenetics of Alzheimer’s disease and related dementias in Latin American countries’ (1R01AG082056-01A1)), the GBHI and the Alzheimerʼs Association (‘Brain health in individuals with exposition to high violence in Colombia’ (GBHI ALZ UK-23-971135)). J.E. acknowledges support from the UKRI AI program and the Engineering and Physical Sciences Research Council, for the CHAI-EPSRC AI hub for Causality in Healthcare AI with real data (grant number EP/Y028856/1). J.D.S. is supported by Program Investissements d’avenir ANR-10-IAIHU-06. M.S.-A. is supported by CONCYTEC (contrato no. PE501082737-2023-PROCIENCIA). L.T.I. is supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, grant number 2022/15880-5). A.T. is supported by Investment for the Future Programme France 2030 (grant no. ANR-18-RHUS-0002) and the France 2030 IHU3 initiative for the Precision and Global Vascular Brain Health Institute (grant ANR-23-IAHU-0001). Y.J. is supported by NIA, NIH P01AG078116 and P30AG072946. (Data curation was partially supported by NIH grant AG000986 and Department of Energy contract DE-AC05-00OR22725.) C.B. is supported by funds of the project ‘Rome Technopole’ from the call Piano Nazionale di Ripresa e Resilienza (PNRR), Mission 4, ‘Istruzione e Ricerca’, Component 2, ‘Investment 1.5’, granted by the European Union, Next Generation EU and the Italian Ministry of University and Research (grant agreement ECS00000024). J.B.S. is supported by CONADI-Universidad Cooperativa de Colombia. D.H. is supported by ANID/FONDECYT Regular number 1231117. O.P. is supported by an NHMRC of Australia program grant (GNT1037746) and a Leadership Fellowship (GNT2008020) and by the Australian Research Council Centre of Excellence in Cognition and its Disorders Memory Program (CE11000102). P.A.V.S. is supported by the National Key R&D Program of China (2024YFE and 0215100). S.G. is supported by the Deutsche Forschungsgemeinschaft (DFG, GE 2835/2–1 and GE 2835/9-1). D.J.S. was supported by the SAMRC. F.M. is supported by Spanish Ministry of Science PID2021-122979OB-C21. M.P. is supported by the IRCCS Istituto Centro San Giovanni di Dio Fatebenefratelli and acknowledges support from the Italian Ministry of Health (Ricerca Corrente). The principal investigator of ARWIBO is G. B. Frisoni, University Hospitals and University of Geneva, Geneva, Switzerland, and IRCCS Fatebenefratelli, the National Centre for Alzheimer’s and Mental Diseases, Brescia, Italy. ARWIBO is the result of efforts of many researchers of IRCCS Fatebenefratelli: G. Binetti, Neurobiology; L. Bocchio-Chiavetto, Neuropharmacology; M. Cotelli, Neuropsychology; C. Minussi, Neurophysiology; M. Gennarelli, Genetic Unit; R. Ghidoni, Proteomics Unit; and D. Moretti and O. Zanetti, Alzheimer’s Unit. Data collection and sharing for ARWIBO was supported by the Italian Ministry of Health under the following grant agreements: Ricerca Corrente IRCCS Fatebenefratelli, Linea di Ricerca 2; Progetto Finalizzato Strategico 2000−2001 ‘Archivio normativo italiano di morfometria cerebrale con risonanza magnetica (età 40+)’; Progetto Finalizzato Strategico 2000−2001 ‘Decadimento cognitivo lieve non dementigeno: stadio preclinico di malattia di Alzheimer e demenza vascolare. Caratterizzazione clinica, strumentale, genetica e neurobiologica e sviluppo di criteri diagnostici utilizzabili nella realtà nazionale’; Progetto Finalizzata 2002 ‘Sviluppo di indicatori di danno cerebrovascolare clinicamente significativo alla risonanza magnetica strutturale’; Progetto Fondazione CARIPLO 2005−2007 ‘Geni di suscettibilità per gli endofenotipi associati a malattie psichiatriche e dementigene’; ‘Fitness and Solidarietà’; and anonymous donors. M.S. is supported by the NIA (grant 1R01AG081305); Davos Alzheimer’s Collaborative grant DAC-GC-22240710.01; the Doha International Family Institute (grant OSRA6-G-202305-00052); and American University in Cairo Research Support Grant 2022. H.B. has received funding from the NHMRC. P.S.S. is supported by an NHMRC of Australia Investigator Grant (RG193540), an NHMRC CRE grant (RG203943) and an NIH grant (R01AG057531-03). The Sydney Memory and Ageing Study has been funded by three NHMRC program grants (ID350833, ID568969 and APP1093083). J.J.M. is supported by the Academy of Medical Sciences (NGR2/1210); the Alliance for Health Policy and Systems Research (2009/32034 and 2012/253750); Bloomberg Philanthropies (grant 46129, via University of North Carolina at Chapel Hill School of Public Health); FONDECYT via CIENCIACTIVA/CONCYTEC, the British Council, the British Embassy and the Newton-Paulet Fund (223-2018 and 224-2018); DFID/MRC/Wellcome Global Health Trials (MR/M007405/1); the Fogarty International Center (R21TW009982, D71TW010877, R21TW011740 and K01TW011478); Grand Challenges Canada (GMH-POC-0335-04); the International Development Research Center Canada (IDRC 106887 and 108167); the Inter-American Institute for Global Change Research (IAI CRN3036); the National Cancer Institute (NCI 1P20CA217231); the National Council for Scientific and Technological Development (CNPq Brasil 408523/2023-9); the NHMRC (2022036, 2022566 and 2044237); the National Heart, Lung, and Blood Institute (NHLBI HHSN268200900033C, 5U01HL114180 and 1UM1HL134590); the NIA (R01AG057531); the National Institute for Health and Care Research (NIHR150261, NIHR150287, NIHR303125 and NIHR306208); the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK K23DK135798); the National Institute of Mental Health (NIMH 1U19MH098780); NSW Health (H23/37663 and DG23/7050); the Medical Research Future Fund (MRRF 204223); the Swiss National Science Foundation (40P740-160366); UKRI BBSRC (BB/T009004/1); UKRI EPSRC (EP/V043102/1); UKRI MRC (MR/P008984/1, MR/P024408/1, MR/P02386X/1, MR/X004163/1 and MR/X020851/1); Wellcome (074833/Z/04/Z, 093541/Z/10/Z, 103994/Z/14/Z, 107435/Z/15/Z, 205177/Z/16/Z, 214185/Z/18/Z and 218743/Z/19/Z); the World Diabetes Foundation (WDF15-1224); and the World Health Organization (2021/1189041 and 2022/1249357). T.E. is supported by the William H. Gates Sr. Fellowship from the Alzheimer’s Disease Data Initiative and Alzheimer Nederland Project WE.03-2024-07. S.B. is supported by the GBHI, the Alzheimer’s Association, Alzheimer’s Society UK and Pilot Awards for Global Brain Health Leaders (grant GBHI ALZ UK- 25-1289623). E.T. is supported by ANID/FONDECYT Regular 1220995 (Chile), ‘Mapping and modeling healthy and pathological aging trajectories’ and FONDECYT Exploración 13240170 ‘Neurovascular pathways in aging and neurodegenerative diseases: integrating cardiovascular health and brain function’. C.D.-A. is supported by ANID/FONDECYT Regular 1210622, ANID/PIA/ANILLOS ACT210096, the Alzheimer’s Association (AARGD-24- 1310017), ANID/FOVI240065 and ANID/Proyecto Exploracion 13240170. A.I. is supported by grants from the ReDLat (supported by the Fogarty International Center, the NIH, NIA (R01 AG057234, R01 AG075775, R01 AG21051, R01 AG083799, CARDS-NIH and R01 AG057234), the Alzheimer’s Association (SG-20-725707), the Rainwater Charitable Foundation, the Bluefield Project to Cure Frontotemporal Dementia and the GHBI); ANID/FONDECYT Regular (1250091, 1210176 and 1220995); ANID/PIA/ANILLOS ACT210096; JPI JPND-Care, DISCeRN 2025: Health and Social Care Research with a Focus on the Moderate and Late Stages of Neurodegenerative Diseases; FONDEF ID20I10152 and ANID/FONDAP 15150012; Wellcome Trust award for BRAIN-CLIMA: Investigating the Combined Impact of Heat and Air Pollution on Blood−Brain Barrier Integrity and Brain Aging in Latin America (335293/Z/25/Z); and CliCBrain (Horizon ID: 101236426; https://doi.org/10.3030/101236426 Marie Skłodowska-Curie Actions). The contents of this publication are solely the responsibility of the authors and do not represent the official views of these institutions. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
We dedicate this work to the memory of our co-authors, Dan J. Stein and Eman Khedr, who passed away in 2025.
Author information
Authors and Affiliations
Contributions
A.I., A.L., S.M. and H.H. were responsible for conceptualization, formal analysis and writing the original draft. All authors contributed to review and editing. A.L., S.M., H.H., P.B., J.C. and S.D.G. had access to the raw data. All authors had access to all preprocessed data in the study, and the corresponding authors had final responsibility for the decision to submit for publication.
Corresponding authors
Ethics declarations
Competing interests
All authors declare no competing interests.
Peer review
Peer review information
Nature Medicine thanks Li Feng, Smadar Shilo and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Ming Yang, in collaboration with the Nature Medicine team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Individual GAMs for structural BAGs – total exposome.
a, Scatterplots show top-ten individual model GAMs between each exposomal factor and structural BAGs. Solid red lines represent the mean GAM fit; shaded bands indicate 95% confidence intervals. P-values reflect the significance of GAM predictive accuracy, assessed via two-sided t-test on the prediction-observation slope in held-out data. All models are FDR P < 0.001. Scatter plots were generated in Python using Matplotlib. Created in BioRender; Migeot, J. (2026) https://biorender.com/y73zai8 (MRI device). Related to Fig. 2.
Extended Data Fig. 2 Individual GAMs for functional BAGs – total exposome.
a, Scatterplots show top-ten individual model GAMs between each exposomal factor and functional BAGs. Solid red lines represent the mean GAM fit; shaded bands indicate 95% confidence intervals. P-values reflect the significance of GAM predictive accuracy, assessed via two-sided t-test on the prediction-observation slope in held-out data. All models are FDR P < 0.001. Scatter plots were generated in Python using Matplotlib. Created in BioRender; Migeot, J. (2026) https://biorender.com/rxp5lws (MRI and EEG device). Related to Fig. 2.
Extended Data Fig. 3 Individual GAMs for combined BAGs – total exposome.
a, Scatterplots show top-ten individual model GAMs between each exposomal factor and combined BAGs. Solid red lines represent the mean GAM fit; shaded bands indicate 95% confidence intervals. P-values reflect the significance of GAM predictive accuracy, assessed via two-sided t-test on the prediction-observation slope in held-out data. All models are FDR P < 0.001. Scatter plots were generated in Python using Matplotlib. Created in BioRender; Migeot, J. (2026) https://biorender.com/rskoaw4 (MRI and EEG device). Related to Fig. 2.
Extended Data Fig. 4 BAGs differences between groups.
a, Violin plots for the distribution of prediction gaps by group. Statistical comparisons were calculated using two-sided subsample permutation testing without multiple comparisons, and with 5,000 algorithm iterations and FDR corrected P values. White dots indicate mean values. Expected significant differences between groups were observed for structural, functional and combined BAGs (healthy controls > MCI > AD > FTLD). **** FDR P < 1E-15; *** FDR P = 9E-04; ** FDR P = 0.002; * FDR P = 0.013. Related to Fig. 2.
Extended Data Fig. 5 Exposome vs clinical condition: odds ratios comparison.
a, Violin plots show the distribution of odds ratios obtained from logistic models using either exposome factors (gray) or clinical condition (purple) as predictors of accelerated aging (binary outcome: accelerated vs. delayed). Panels depict results for (a) total, (b) physical, and (c) social exposomes across structural, functional, and combined modalities (n = 100 per combination). Across all domains, exposome-based models yielded significantly higher odds ratios than clinical-condition-based models (two-sided permutation tests; 5,000 permutations; Bonf P < 1E-15; Cohen’s d = 3.76–17.6), indicating that exposomal burden exerts a stronger and more consistent influence on the likelihood of accelerated brain aging. Box centers represent median values; white lines denote means; box limits indicate interquartile ranges (Q1–Q3), and whiskers represent minimum and maximum observed values. Related to Fig. 3.
Extended Data Fig. 6 Variance inflation factors for exposome and demographic/cognitive variables.
Variance inflation factor (VIF) values assessing multicollinearity among covariates in (a) the in-sample dataset and (b) the SES subsample. Each panel displays VIFs for exposome indices (total, social, and physical) across structural, functional, and combined brain age gap models. Variables include exposome, sex, years of education, cognitive score, age, and individual socioeconomic status (SES; in panel b only). All VIF values were below 1.82, confirming minimal multicollinearity among predictors. Related to Fig. 3.
Extended Data Fig. 7 Sensitivity analyses: Robustness across individual variables and methodological adjustments.
(a) SHAP results with imputed education and cognition. SHAP sensitivity analyses where re-run with imputed values for education (16,4% imputed) and cognition (36,7% imputed). GDP data were complete except for Cuba (see missingness details per modality in Supplementary Table 42 and handling strategy and summary Supplementary Table 43). Results remained consistent with non-imputed analyses. (b) Participant demographics and country level GDP effects. Individual SES effects on brain clocks were tested with XGBoost models, including other individual (sex and cognition) and aggregate (country-level GDP) variables. Lower SES was associated with worse outcomes, along with lower cognitive scores, and female sex (T1 model), and lower GDP and cognitive scores (functional model). Shapley additive explanations (SHAP) values indicate the direction and magnitude of each feature’s contribution. Violin plots show SHAP distributions; dots represent participants. (c) MetaGAMs covariation. The effects of total exposome metaGAMs on accelerated brain aging in the subsample with SES remained after controlling for covariates. Violin plots show bootstrapped effect sizes; dots are samples (n = 5,000), red lines are the mean ΔAIC, black lines indicate 95% confidence intervals. (d) DeltaAIC differences between metaGAMs derived from uncorrected and age-bias corrected BAGs. Violin plots showing the distribution of bootstrap metaGAM ΔAIC values (n = 5,000) by BAG condition in the total sample. Statistical comparisons were calculated using two-sided subsample permutation testing without multiple comparisons, and with 5,000 iterations and FDR P values. Data are presented as mean ΔAIC values ± 95% confidence intervals (red lines). No significant differences between BAG conditions were observed for structural, functional and combined BAGs across total, physical and social exposomes (all FDR P > 0.001), supporting the stability of exposome-related findings regardless of age-bias correction. Related to Fig. 2.
Extended Data Fig. 8 Sensitivity analyses: Robustness across health, disease, and imaging quality.
(a) Brain features in healthy controls BAG models. Top-ten brain areas contributing to healthy controls BAGs in structural (n = 7,627, R² = 0.75, F² = 2.94, MDE = 0.02) and functional (n = 6,692, R² = 0.41, F² = 0.69, MDE = -0.04; all P < 0.001) models. Feature importance was derived from SHAP (Shapley additive explanations). Data are presented as mean SHAP values (red line) with 95% confidence intervals (black error lines), derived across n = 10 outer fold samples (dots). (b) DeltaAIC differences between controls and patients metaGAMs. Violin plots showing the distribution of bootstrap metaGAM’s ΔAIC by subsample. Statistical comparisons were calculated using two-sided subsample permutation testing without multiple comparisons and with 5,000 algorithm iterations and FDR P values. Expected significant differences between subsamples were observed for structural, functional and combined BAGs (Healthy controls > patients) across total, physical and social exposomes (all FDR P < 0.001). (c) ODQ in clinical conditions. Association between ODQ and BAGs in structural and functional models. Linear regressions used permutation testing (10,000 repetitions) to assess significance. BAG values were randomly permuted to generate a null distribution of regression slopes for FDR P value calculations. No significant associations were found. (d) Disease severity and BAGs in patients with dementia. Linear regressions using permutation testing (100,000 iterations) were run to target the association between CDR scores and structural and functional BAG values across patients. Each line denotes the fitted linear trend within patient subgroups (AD, FTLD) and all patients together. In both modalities, higher CDR scores were associated with increased BAGs across conditions. Kernel density contours indicate the distribution of BAG values across the CDR spectrum. * indicates P FDR < 0.001. Related to Fig. 2 and Fig. 4.
Extended Data Fig. 9 Separate BAGs for rsfMRI and rsM/EEG.
a, Scatterplots show model accuracy for predicted brain age versus chronological age using functional rsfMRI (left panel) and functional rsEEG/MEG (right panel) in-sample data. Brain plots indicate the top-10 most important brain features ranked by Shapley additive explanations (SHAP) values for each model. Reported P-values (P < 1E-15) are two-sided and derived from a t-test on the regression slope coefficient; no adjustments for multiple comparisons were applied. Related to Fig. 2.
Extended Data Fig. 10 Sex-specific associations between gender inequality and brain aging.
a, Violin plots display the distribution of correlation coefficients (r values) between brain age gaps (BAGs) and gender inequality measures (gender inequality index and inverse gender equality (rights)) computed separately for females and males. Each panel represents the correlation pattern across structural, functional, and combined BAGs. Sex differences were assessed using two-sided t-tests on correlation coefficients (n = 100 per sex), without adjustment for multiple comparisons. Statistical tests (shown above each plot) indicate significantly stronger positive associations in females (all P < 0.01), suggesting that higher gender inequality is more tightly linked to accelerated brain aging in women than in men. Related to Fig. 2.
Supplementary information
Supplementary information (download PDF )
Supplementary Figs. 1 and 2 and Supplementary Tables 1−46
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article
Cite this article
Legaz, A., Moguilner, S., Barttfeld, P. et al. The exposome of brain aging across 34 countries. Nat Med (2026). https://doi.org/10.1038/s41591-026-04302-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41591-026-04302-z
Facts Only
Researchers: University of Barcelona and collaborators from 34 countries
Study type: Analysis of the exposome's effect on brain aging using a dataset of over 70,000 older adults
Factors investigated: Environmental, socio-economic, and lifestyle factors (the exposome)
Key findings: Gender inequality, pollution, diet, socio-economic status significantly impact brain health in later life
Executive Summary
Full Take
This study sheds light on the complex interplay between various environmental, social, and personal factors contributing to cognitive decline in older adults. By employing a multifaceted approach and analyzing data from a large, diverse sample, it challenges us to reconsider traditional notions of brain health as solely determined by genetics or age. However, the research also raises important questions about the ethical implications of using comprehensive datasets that may include sensitive information, particularly in cross-cultural contexts. Furthermore, the study underscores the need for global cooperation to address factors such as pollution and gender inequality that affect brain health on a broad scale.
Patterns detected: ARC-0024 Ambiguity (the study's findings may be interpreted differently depending on the reader's perspective)